本章節介紹身體各項知覺。
深度知覺
编辑視知覺給我們提供一個重要的能力:即使視網膜成像是二維的,我們仍能看到三維世界。差別在於所接收到的訊息深度不同,分為單眼線索和雙眼線索。單眼線索是只需要一隻眼睛就能感受的線索;雙眼線索是需要同時用兩隻眼睛才能感受到的線索。若我們將一隻眼睛閉起來,我們還是多少能夠感知一些深度的訊息,這些訊息被稱之為單眼深度線索(monocular depth cues),或者是圖畫線索(pictorial cues),因為這種線索利用了藝術家在繪製圖畫時會用到的技巧,如透視線。
部分人患有立體盲,不能利用視網膜的成像差異適當估計距離。最常見的立體盲是斜視,兩眼從不同的方向看向物體。這是一種嚴重的缺陷,可導致疊像的產生,或即使雙眼都睜開時,卻只靠一隻眼睛判斷距離。
靜態線索
编辑- 雙眼深度線索(binocular depth cues)
我們的雙眼藉由兩眼視網膜呈像的像差(retinal disparity)以及雙眼聚合(convergence)來提供深度訊息。
- 視網膜像差:大腦藉由兩眼間距所造成其所接收外界影像不同所產生的「像差」,來解讀物品在我們這個三元空間中的深度。例如,若有A、B兩物體,此兩物體在兩眼中所見的距離可能因為左右眼視角的不同而有大小的差異,若此AB間的距離在兩眼中看起來類似,可能代表AB相距不遠;反之,若AB間距離在兩眼中有較大的差異,可能代表AB有距離上的不同。
- 雙眼聚合(convergence):講述當我們凝視物體時,雙眼瞳孔轉向內側的情況(有點像是輕微鬥雞眼的狀況)。若物體離我們十分近,則眼睛需要內轉較多才能聚焦在物體上,因此大腦利用來自眼球肌肉的訊息來了解近距離物體的遠近。
我們如何把兩個不同圖像組合或匹配成單一的知覺?按照馬爾和葡基耶(Marr & Poggio)的理論,有三條主要原則
- 兼容性原則:為了使兩個視網膜成像相配,這兩個部分要在某地方是兼容的或相似的,例如顏色和形狀。
- 連續性原則:如果兩個視網膜像之間的差異與已經建立的鄰近視網膜像之間的差異類似,這兩個視網膜像更容易匹配。
- 唯一性原則:一個視網膜像的某一個部分僅與另一個視網膜像的部分相配。
- 單眼深度線索(monocular depth cues):
- 質地梯度(texture gradient)
- 距離近的物體將會有較多的細節資訊,其內容是指藉由觀察遞減的紋理來了解距離,簡單而言就是因為越往深處,物體質地密度在視覺上會變得越大(這是另一個距離、大小關係所造成的結果),因此可以藉由這種「材質紋理」來了解三次元空間中的距離。
- 遮擋(Interposition)
- 對於深度視覺來說,最重要的單眼線索之一是遮擋。當一個近的物體部分的遮住了遠處物體時,就出現了遮擋。卡尼撒(Kanizsa)的錯覺方塊支持了這一個線索存在的想法。它的錯覺方塊是四顆黑球擺在正方形的四個頂點,然後內側個被挖去四分之一圓,這個時候,視覺系統在中間產生了一個白色的方塊,遮擋在黑球前面,使得圓的缺失部分變的合情合理。
- 直線透視(linear perspective)
- 最簡單的例子就是無限延伸的鐵軌在視野中的最盡頭會似乎會匯合到一個點上,鐵軌應該是由兩條平行線所組成,可因為龐氏錯覺(Ponzo illusion),所以比較接近的物體會看起來較長、而比較遠的物體則是看起來較小、較短;
- 熟悉大小(Familiar size)
- 另一個重要的深度線索是熟悉大小。如果我們熟知一個物體的大小,那麼,我們就可以用它在視網膜上的大小計算出距離。
- 大氣透視法(Atmospheric perspective)
- 大氣透視,又稱空氣透視或空氣遠近法,是由於受到空氣微粒的影響,使得近、中、遠距離的物體會因為光學作用而產生不同程度的顏色變化,人們看到近處的景物比遠處的景物濃重、色彩飽滿、清晰度高。受大氣透視影響越嚴重對比度就會越低,所以也可以說越遠的東西失去越多細節表現。正中午的大氣透視最弱,清晨及夕陽的斜角光線會讓透視感較為顯著,而濃霧或嚴重空氣汙染會產生更強烈的大氣透視效果,例如”伸手不見五指”的情況通常就是濃霧所造成的。這廣泛運用在繪畫和攝影技巧之中。
動態線索(Motion cues)
编辑- 屬於單眼探索
- 運動視差(motion parallax)這種線索是物體在視網膜成像上的運動。一般來說,對於離觀察者較近的運動物體,其視網膜成像的運動量比離觀察者遠的物體更大,還有一個現象與運動視差有關。如果你在行駛很快的汽車裡,從車窗向外看,離你很遠的物體看起來似乎是跟汽車同向移動,而較近的物體似乎在向相反方向移動。
視錯覺
编辑視錯覺(Optical illusion)是指透過幾何排列、視覺成像規律等手段,製作有「視覺欺騙」成分的圖像引起的視覺上的錯覺。視錯覺一般被分為:
- 圖像本身的構造導致的幾何學錯覺
- 由感覺器官引起的生理錯覺
- 心理原因導致的認知錯覺
其中特別是關於幾何學的錯覺,以其種類多而廣為人知。
- 幾何學錯覺:視覺上的大小、長度、面積、方向、角度等幾何構成,和實際上測得的數字有明顯差別的錯覺,稱為幾何學錯覺,以下列出幾個較為著名的幾何學錯覺:
-
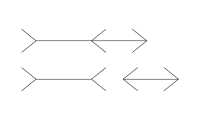 (繆莱二氏錯覺):由德國社會學家弗朗茨·米勒-萊爾於1889年發現,是指兩條等長平行線段,兩端箭頭向外的線段比兩端箭頭向內的線段看上去更長的現象。
(繆莱二氏錯覺):由德國社會學家弗朗茨·米勒-萊爾於1889年發現,是指兩條等長平行線段,兩端箭頭向外的線段比兩端箭頭向內的線段看上去更長的現象。 -
 艾賓浩斯錯覺:是一種對相對大小感知的一種視錯覺,以發現者,德國心理學家赫爾曼·艾賓豪斯(1850-1909)的名字命名,當兩個完全相同大小的圓放置在一張圖上,其中一個圍繞較大的圓,另一個圍繞較小的圓,而圍繞大圓的圓看起來會比圍繞小圓的圓要小。最近的研究表明,影響艾賓浩斯錯覺感知的另外兩個關鍵因素是週圍圓圈與中心圓的距離和圓環的完整性。
艾賓浩斯錯覺:是一種對相對大小感知的一種視錯覺,以發現者,德國心理學家赫爾曼·艾賓豪斯(1850-1909)的名字命名,當兩個完全相同大小的圓放置在一張圖上,其中一個圍繞較大的圓,另一個圍繞較小的圓,而圍繞大圓的圓看起來會比圍繞小圓的圓要小。最近的研究表明,影響艾賓浩斯錯覺感知的另外兩個關鍵因素是週圍圓圈與中心圓的距離和圓環的完整性。 -
 黑林錯覺:於1861年由德國生理學家埃瓦爾德·黑林(Ewald Hering)發現,指兩條平行直線在徑向背景(如自行車輻條)前時,看起來會向外彎曲的現象,此種錯覺亦稱為彎曲錯覺。
黑林錯覺:於1861年由德國生理學家埃瓦爾德·黑林(Ewald Hering)發現,指兩條平行直線在徑向背景(如自行車輻條)前時,看起來會向外彎曲的現象,此種錯覺亦稱為彎曲錯覺。 -
 加斯特羅圖形:由美國心理學家約瑟夫·賈斯特羅(Joseph Jastrow)發現於1892年,兩扇形雖然大小形狀完全相同,但是下方的扇型看似更大。
加斯特羅圖形:由美國心理學家約瑟夫·賈斯特羅(Joseph Jastrow)發現於1892年,兩扇形雖然大小形狀完全相同,但是下方的扇型看似更大。




- 生理錯覺:主要是因為人體的視覺適應現象,人的感覺器官在接受過久的刺激後會鈍化,也就造成了補色及視覺暫留的生理錯覺。當視網膜上的細胞受某種色光刺激後,會對該色產生疲勞,所以在視線離開該色後,該部分的細胞暫無法作用,而未受刺激的另一部分細胞開始活動,因而產生另一種視感,也就是補色的殘像。此外,過視覺疲勞而產生的視覺暫留現象就是現今動畫的原理。以下列出幾個著名的生理錯覺例子:
-
 赫曼方格錯覺:在黑色背景上的白色(或淺色)網格的交叉點上,可以看到 "幽靈般"的灰色斑點,而當直接看向一個交叉點時,灰色斑點會消失。
赫曼方格錯覺:在黑色背景上的白色(或淺色)網格的交叉點上,可以看到 "幽靈般"的灰色斑點,而當直接看向一個交叉點時,灰色斑點會消失。 -
 馬赫帶:沿著相鄰的灰色陰影之間的邊界,側向抑制使較暗的區域會假性地顯得更暗,而較亮的區域則假性地顯得更亮。
馬赫帶:沿著相鄰的灰色陰影之間的邊界,側向抑制使較暗的區域會假性地顯得更暗,而較亮的區域則假性地顯得更亮。


- 認知錯覺:這主要來自於人類的知覺恆常性屬於認知心理學的討論範圍,如ASCII立體圖(ASCII stereogram)、奈克方塊(Necker cube)、圖地反轉(Negative space)及鴨兔錯覺(Rabbit–duck illusion)等
前庭訊息的處理
编辑前庭系統vestibular system是負責人體自身平衡感和空間感的感覺系統,對於人的運動和平衡能力有關鍵性的作用。而前庭訊息處理使眼、頭、身體能夠作出補償的運動以對抗各種機械運動。它的運作主要是由從內耳的前庭感受器出來的神經纖維,進入腦幹的低級中樞,與前庭核(vestibular nuclei)的神經元形成突觸連結,某些前庭神經纖維則會繞過前庭核直接進入小腦(重要的運動中樞)。
前庭也常常使人暈船或是暈機,這是身體的加速度感受器引起前庭系統興奮所導致的現象,我們稱之為暈症(motion sickness)。熱也能產生相同的效果,比如說在耳朵裡灌溫水,會引起內耳的淋巴液運動,產生機械刺激導致頭暈。因藥物或抗生素中毒而傷害前庭系統的病人不會有暈症。
嗅覺訊息的處理
编辑嗅覺黏膜上擁有數百萬個嗅覺細胞,各自只表現一種嗅覺受體;同時,單一種氣味分子,可以活化不只一種嗅覺受體。因此,任何一種嗅覺,都是由不同數量及組合的嗅覺細胞,受到不同程度的活化或抑制後,將訊息傳入嗅覺中樞,再經過大腦解碼下的產物,非單純一對一的關係。
嗅覺細胞為少數的雙極神經元(bipolar neuron,僅有一個樹突和一個軸突的神經元),其樹突端往下伸入鼻腔,接收吸入的氣味分子;軸突端則向上穿過頭骨,進入嗅球(olfactory bulb)。帶有相同受體的嗅覺細胞軸突,會在嗅球中匯集成一個嗅小球(glomerulus),每個嗅小球有25至50個來自僧帽細胞(mitral cell)的樹突進駐。因此,嗅小球是嗅覺訊息的第一個整合中心。僧帽細胞將訊息進行區分及放大之後,其軸突形成嗅神經束(olfactory tract),傳送至嗅覺皮質作進一步的處理。
嗅覺是所有感覺系統當中,唯一不需經過脊髓或間腦的轉接,就能直接投射至前腦的感覺系統;其投射也如其他的感覺系統具地域性分布的特性,在大腦嗅覺皮質上形成地圖般的構造。此外,嗅覺訊息還可以送往邊緣系統(limbic system);由於邊緣系統是負責情緒、記憶及行為的腦區,因此也可以解釋嗅覺具有引發強烈情緒及記憶的作用。
在近年對於鼻腔的嗅粘膜和嗅球的解剖研究發現,這些部位在反映個氣味上有區域性的劃分,也就是說一定的區域對於一定的氣味特別敏感。然而,確定各氣味的感受區極端困難,因為劃分氣味本身就是一個難題,至今仍未找到氣味分子的特定化學成分。因此,研究嗅覺的訊息處理遇上了阻礙,因為我們發現分子的大小和形狀這類的化學特性和嗅覺系統的訊息編碼過程沒有太大的關係。
味覺訊息的處理
编辑味覺訊息經由三條腦神經送入腦中,分別是顏面神經中的谷索神經(chorda tympani),傳送舌頭前三分之一的味覺感受氣的訊息;舌咽神經(glossopharyngeal nerve)傳送舌頭後三分之一的味覺感受氣的訊息;迷走神經(vagus nerve)傳送軟顎和咽部的味覺感受氣的訊息。這些傳送味覺訊息的神經有些與延髓中的孤立核(solitary nucleus)的神經元形成突觸連結;有些與橋腦中的味覺中樞的神經元形成突觸連結。突觸神經元的軸突將味覺訊息傳到視丘,再由視丘神經元傳到大腦皮質的味覺區。
味覺的訊息處理較少被研究,除了是因為人們覺得味覺比較簡單外,味覺對現代人的生存已無太大影響。神經學家在味覺主要進行的細胞放電紀錄主要是分析鹹、酸、甜、苦的訊息編碼。一派學者認為,一種細胞只反應一種味覺刺激;另一派認為味道是由眾多細胞不同的興奮模式整合所決定,稱為交叉纖維模式理論(crossfiber patterning theory)。 值得注意的是,過去有學者認為舌頭的特定位置會感覺到不同的味道,然而實驗已證明味覺和味蕾的位置並沒有明顯的對應,也就是舌頭上的任意位置都能感覺到不同的味道。
聽覺訊息的處理
编辑腦的聽覺構造與通路
编辑聲音的訊號在進入耳朵後,將會經過外耳(Outer ear)、中耳(Middle ear)以及內耳(Inner ear),最後傳達到內耳的纖毛細胞,以下將逐一介紹耳朵的構造。
- 外耳(Outer ear)
- 外耳包含了耳廓(Auricle)與外耳道(External acoustic),主要功能是接收外來的聲能,將其傳至中耳的鼓膜。耳殼就是平時我們肉眼可見的部分,可以負責收集聲波,它與與外耳道口是由結締組織相連;而外耳道則是從耳廓延伸到鼓膜,由軟骨及骨骼組成,外耳道也能藉由共振(resonance)的作用,來增加聲音強度,使我們能對於聲音頻率的範圍更加敏銳
- 中耳(Middle ear)
- 中耳包括了鼓膜(Tympanic membrane)、鎚骨(Malleus)、砧骨(Incus)還有鐙骨(Stapes),主要的機能就是放大聲音訊號。鼓膜是耳朵的重要構造,它是一層較硬的膜狀構造,其主要負責的工作是以振動方式傳遞聲音,並將之傳給中耳的聽小骨(由鎚骨、砧骨與鐙骨組成),鼓膜破損的話也有可能導致傳導性聽力損失。鼓膜的兩側分別為外耳道及鎚骨,鎚骨另一側又與砧骨相接,而砧骨的另一邊也與鐙骨相連,最後傳至卵圓窗,而它連接著前庭。而這些構造提供了兩種增強音訊的機制。首先,鼓膜到卵圓窗的面積是由大到小,能有效提高單位面積壓力;再來,聽小骨之間能利用槓桿原理將聲波放大,來防止聲波因為從外耳、中耳裡的空氣到內耳的耳蝸內液體(淋巴液),而造成能量的流失。而聽小骨中的砧骨也與一些骨骼肌相連,而這份功能能藉由肌肉的反射機制來降緩振動,以保護耳朵、聽力。
- 內耳(Inner ear)
- 內耳主要是由前庭系統(Vestibular system)與耳蝸(Cochlea)組成,能將機械波振動轉換成電訊號,再傳到大腦中的聽覺皮質,而其中前庭神經負責傳送平衡覺信息,耳蝸神經負責傳送聽覺信息。
而每一根聽神經纖維再進入腦幹之後會分為兩大枝,一枝到耳蝸前核(ventral cochlear nuclei),另一枝到耳蝸背核(dorsal cochlear nuclei)。從這兩個核出來的纖維,一枝到上橄欖核群(superior olivary complex,接受右左兩耳蝸發出的纖維,為聽覺系統第一級雙耳輸入交互作用的地方,為聽覺定位機制的重要部位),其他平行的路線匯聚在下丘核(medial geniculate nucleus),到中腦的聽覺中樞。
聽覺系統和其他系統一樣,具有上行和下行通路,且比其他感覺系統更加明顯。下列傳出纖維多來自橄欖核,它們在耳蝸之中分枝很多,與每一個毛細胞的基底部形成突觸連接。
音調的辨別
编辑我們可以聽到的波頻為 20-20000Hz,在這個範圍內我們可以辨別極小的頻率差別,在 2000Hz 以內可察覺的差別約為 2Hz。音調(pitch)和頻率(frequency)不同,音調指的是感覺經驗,而頻率為聲音的物理特性。頻率不是決定音調的唯一因素,而音調的改變和頻率的改變也不完全一致。辨別頻率有三種理論:
- 地點理論(place theory)
- 此理論認為音調知覺決定於基底膜的波動最大的地點(「地點」應包含中樞接受興奮的神經元地點),也就是說特定的神經元反應特定的刺激頻率。
- 齊發理論(volley theory)
- 此理論強調聲音刺激的頻率和神經放電的模式或時間有關係(例如:500Hz 的純音應該代表每秒 500 次的神經衝動),意味著眾多神經元同時反映一個聲音的刺激。單個神經元的放電模式會反映刺激的頻率,刺激頻率的改變也會使放電模式改變,而頻率和神經衝動的一致性若由數根纖維來達成會比由單根纖維達成更為精準,因此用「齊放」來形容。
- 雙重理論(duplex theory)
- 結合了地點理論和齊放理論。我們觀察到基底膜的波動最大的地點與頻率有關,刺激頻率變了,基底膜的波動最大的地點也會改變。例如,人們觀察到在某些頻率的聲音刺激時,耳蝸內可以完成一種傅立葉分析;不同的頻率表現在基底膜不同的地段上。基底膜上的表面隨著演化的過程中的加長,毛細胞、神經纖維的增加,都使得辨認聽覺頻率更加的精確。
在腦中,各級神經元各有其最敏感的頻率。我們可以用神經元放電的時間模式來作為音頻訊息的編碼過程,其中包括以興奮細胞的分佈來編碼(又稱地址編碼),以及從神經到聽覺皮質的各級中樞神經元放電時間模式來編碼。
聲音的定位
编辑聽覺定位的原理為,兩耳之間聲音強度的不同以及聲音到達兩耳的時間不同的情況下,兩耳處理系統的結果。對於低頻的聲音定位,最重要的線索是聲音到達兩耳的時間差;而對於高頻的聲音定位,主要是靠兩耳之間的強度差。在最好的條件下,一個人判斷聲源方位可以達到只有一度的誤差。 人類兩耳在聽到聲音時,聲源位置的察覺可能是由中樞內許多細胞的特殊的空間和時間模式所產生,也可能是更高級中樞的某種處理結果。
在分析貓的皮質神經細胞反應時,沒有發現聽覺空間的特徵偵察器;但在貓頭鷹這類靠精準聽覺定位系統來獵捕動物的鳥類腦中,在相當於下丘核的部位,發現了某些細胞的排列很像空間中的球體代表。每一個細胞都具有空間的特異性,具有像小圓椎體的感受器,圓錐尖端都輻合於貓頭鷹的頭部,每一個細胞只對於自己圓錐感受器的聲源刺激敏感。在貓頭鷹的聽覺頂蓋部(optic tectum),聽覺和視覺的感覺系統互相符合,且所有的聽覺細胞對於空間方位都相當敏感,給予確認獵物位置的雙重訊號。此為貓頭鷹生存所需,亦為天擇的產物。
聽覺雙重理論 Duplex Theory
编辑1907年,雷利勳爵(Lord Rayleigh)受到音叉產生的單音激發,在無耳廓人頭模型上研究了側向聲音定位理論。雷利提出聽覺跨耳線的差異,即所謂的雙重理論。 由於人的耳朵位於頭部的兩側,在空間中具有不同的坐標。由於聲源與耳朵之間的距離不同,兩耳的聲音信號之間存在時間差和強度差。這些差異被稱為聽覺跨耳時間差(ITD)和聽覺跨耳強度差(IID)。 雷利發現: 對於低於800 Hz的聲音頻率,頭部尺寸(即雙耳間距21.5cm)小於聲波的半波長。因此,聽覺系統可以在兩耳之間精確地確定相位延遲,而不會混淆。在這個頻率範圍內,聽覺跨耳強度(ITD)差非常低,特別是低於約200 Hz,因此僅基於強度差異從而精確評估音源輸入方向幾乎不可能。
但對於高於1600 Hz的頻率,頭部尺寸大於聲波的波長。在這些頻率上,僅基於耳間相位的確定輸入方向是不明確的。但是,隨著耳間強度差異變大,聽覺系統會針對這些強度差異進行評估。此外,兩耳之間的延遲仍然可通過一些相位差異和延遲的組合檢測到,這些在高頻率聲音時更為明顯;也就是說,如果有高頻聲音出現,則此開始的延遲可以用於確定相應聲源的輸入方向。
聽覺障礙及其康復
编辑定義
编辑聽覺障礙,係指聽覺能力受到損失,其表現可能包含無法經由聽覺接收聲音,或能聽見聲音但無法辨識聲音內容、意義。
在定義上,聽覺障礙 (Hearing loss) 及聾 (Deafness) 有所差異,參考《身心障礙者教育法案》 (Individuals with Disabilities Education. Act, IDEA) 對二者在孩童身上的分別定義:聽力障礙被定義為「聽力損傷,無論是永久性的還是波動性的,都會對兒童的教育表現產生不利影響」。耳聾被定義為「嚴重的聽力障礙,導致兒童在透過聽力(無論有或沒有放大)處理語言訊息時受到損害」。
聽覺障礙分級
编辑世界衛生組織 (World Health Organization, WHO) 依據純音聽力檢查得出的結果(聽力閾值),將聽力障礙分為五級。而通常的「聽力障礙」多指兩耳聽力皆受損,其分級則依優耳聽覺分級為主。
| 聽力障礙等級 | 相應聽力測量ISO值 | 聽力表現 | 建議 |
|---|---|---|---|
| 0:無損傷 | ≤25dB | 沒有或非常輕微的聽力問題。能夠聽到竊竊私語。 | 無。 |
| 1:輕微損傷 | 26-40dB | 能夠在 1 m 處聽到並重複以正常聲音說出的單字。 | 建議諮詢;可能需要助聽器。 |
| 2:中度損傷 | 41-60dB | 能夠在 1 m 處聽到並使用提高的聲音重複單字。 | 通常建議使用助聽器。 |
| 3:嚴重損傷 | 61-80dB | 對著優耳喊叫時能夠聽到一些話語。 | 需要助聽器。如果沒有助聽器,應教導唇讀。 |
| 4:極嚴重損傷 | ≥81dB | 甚至無法聽到並理解大聲喊叫的聲音。 | 助聽器可能有助於理解單字。需要額外的復健治療。唇讀和有時手語必不可少。 |
聽覺障礙的類型
编辑- 傳達聾——外耳和中耳的病變有關的失聽
- 傳導聾涉及了使耳蝸興奮的機械裝置。可能是因為耳垢度塞住了外耳道、中耳炎或耳骨的錯位妨礙運動等等。一般來說,中耳功能障礙造成的聽力障礙或降低對一切頻率的敏感性,且對高頻率的敏感性喪失較多。
- 感覺神經聾——因耳蝸或聽神經的受損產生的聽覺障礙
- 感覺神經聾可能是因為強噪音、藥物、感染、疾病、顳骨外商、顯管障礙、衰老等等因素,這些情況都可能對空氣傳導、骨頭傳導聲波造成障礙,使完全喪失聽力。
- 藥物引起的聽力喪失多是受藥物毒性的影響。比如說鏈黴素(Streptomycin)、卡那黴素(Kanamycin)、慶大黴素(Gentamicin)雖然對結核病有很好的療效,但其中的耳中毒性質(ototoxic properties)會使病人的耳蝸和前庭受到傷害。利用組織學檢查發現,鏈黴素會破壞基底膜底部(靠近卵圓窗的部分)以及毛細胞,使病人喪失高頻率聽覺,甚至完全喪失聽覺且不能恢復。
- 噪音引起的聽力喪失主要是因為內耳機制受到破壞。若內耳組織發生變化主要是毛細胞被破壞,且外邊的毛細胞比內邊的毛細胞更容易受到影響;若是長時間處於高強度的噪音下,整個柯氏器都會被破壞。有時局部傷害就算沒有另外的受傷也會擴散。
- 中樞聾——由於中樞的聽覺通路(腦幹、視丘、大腦皮質)的損傷產生的聽覺障礙
- 中樞聾往往是由腦傷所引起,但這種聾較少,因為很少有腦的兩側同時受傷。
聽覺障礙的類型 受損部位 受損的原因 傳達聾 外耳和中耳 耳垢堵塞、中耳炎、耳骨的錯位妨礙運動 感覺神經聾 耳蝸或聽神經 自然老年退化、高噪音環境、病菌感染 中樞聾 中樞的聽覺通路 由腦傷所引起
- 治療耳蝸性聾的方法
- 藉由用電流刺激聽神經的方式(因為毛細胞遭到破壞,但聽神經仍完好)來使之興奮,而此興奮傳入腦內後可以喚起聽覺。最理想的方式是利用擴音器控制電刺激,以此模擬毛細胞產生的興奮來誘發聽神經的興奮,雖然不能被稱作治療,但能讓病患聽到人們的說話聲。
<補充>音樂知覺
编辑音樂和語言一樣,都在時間結構上有明顯的變化,但語言有絕對或約定成俗的意義,音樂少有此特性。音樂演奏與欣賞除依賴顳葉聽覺皮層外,也依賴其它腦區,例如Premotor及Supplementarymotor cortices。聆聽音樂時運動皮質的活動使音樂與舞蹈在人類文明中有緊密關係;例如人接受簡單鍵盤音樂之演奏訓練後,對其播放該音樂,不僅聽覺皮質興奮,某些運動皮質亦有活動。一個音樂家演奏熟悉的曲子(但聽不見)與聽該曲調(不演奏),同樣都會興奮聽覺皮質,與運動皮質的某些區域。
音樂知覺與產生:音樂是一個複雜的過程,涉及許多不同的時間歷程,也會激起相對應的神經反應,這些反應的產生都有一定的時間。音樂經常激起情緒反應。音樂知覺也常引起運動反應或想要動的意向,尤其是節奏明快的打擊音樂。
音樂知覺—和語文的關係:音樂知覺是人類相當獨特的能力。和語言一樣,音樂知覺也不易找到相對應的基本單位。有一些研究探討欣賞音樂合理解語言是否會動用同一腦區。例如:Musical structure與文法知識會啟動相同的區域-布若卡氏區(Broca's area)。Musical meaning與語意結構會啟動相同的區域:韋尼克區(Wernicke's Area)。音樂知覺與語言知覺的神經機制有相似也有不相似的地方。
旋律記憶:(Peretz, 1996年之研究):患者Patient C.N.喪失對旋律的記憶,但對音調的知覺正常,同時對於語言辨識也正常。
Semantic dementia(Hseih et al. 2011):右側前顳葉受損,其知識記憶受損,連帶對熟悉歌曲的旋律也受損,所以旋律可能為知識記憶一種,相對的阿茲海默症患者的損害不大。
節奏:fMRI研究顯示聽正常節奏相較於不正常節奏會激發運動與小腦系統的活化。隨著節奏打拍子相較於找出節奏會激發基底核。
有失語症家族成員音調知覺正常,但是節奏知覺受損。前者與右腦有關而後者與左腦語言有關。
先天性音樂失能(congenital amusia):一種音樂處理能力受損的神經發育障礙,影響個體分辨音樂的能力,包括音調的辨識、音樂節奏、旋律以及和諧與否的感知。這種情況並不影響智力或一般聽力,也與學習障礙或聽覺問題無關。患有先天性音樂失調症的人們可能無法辨認旋律的起伏,或者無法分辨不同音樂作品或音調之間的細微差別。以實例來說,他們在唱歌或演奏樂器時可能無法分辨音准。這種失調症並非由缺乏音樂教育或文化背景的差異所引起,而是大腦處理音樂信息的方式有所不同,其右側聽覺區與額下回(Inferior frontal gyrus, IFG)可能出現了問題。值得注意的是,雖然先天性音樂失調症患者在音樂音調辨識上有明顯障礙,但他們處理語言中音調的能力通常不受太大影響,特別是在使用自己的母語時。揭示了人類大腦處理音樂和語言的複雜性和部分獨立的機制。 可能的治療方法: 音樂治療:對於先天性音樂失能患者,音樂治療可以幫助他們加強對音樂的理解和感知能力。這種治療通常由訓練有素的音樂治療師來進行,並可以根據患者的具體情況進行個性化的治療計劃。
聽覺訓練:透過聽覺訓練,患者可以逐步提高對音樂元素(如音調、節奏、旋律等)的識別能力。這種訓練可以通過聽音樂練習、聽力遊戲和其他聽覺認知任務來實現。
認知行為療法:認知行為療法旨在通過改變患者的思維模式和行為來改善其心理健康狀態。對於先天性音樂失能患者,這種療法可以幫助他們處理與音樂相關的情感和挑戰,並學習應對這些情感的有效方法。
神經反饋訓練:神經反饋訓練通常使用生物反饋技術,通過監測患者的生理指標(如腦電圖、心率等)來幫助他們學習調節自己的生理過程。對於先天性音樂失能患者,神經反饋訓練可以幫助他們改善與音樂相關的大腦活動模式,從而提高對音樂的感知和理解能力。
這些治療方法可以單獨應用,也可以結合使用,根據患者的具體情況和需求進行個性化的治療計劃。此外,治療過程中的積極參與和長期執行是取得良好治療效果的重要因素。
- 音樂知覺—音色與情緒:
- 音色(Peretz & Coltheart 模型所沒有處理的):不同樂器演奏同一音調其感覺不同,這來自每個樂器對每個音符的起始過程不一,以及所伴隨的不同泛音(harmonics)強度不同。Samson & Zatorre(1994):右側顳葉損傷破壞音色知覺甚於音調。
- 情緒:音樂會引發情緒反應。節奏快的較節奏慢的音樂聽起來快樂,大調快樂興奮,小調哀傷沈痛。偏離預期的音符引發驚愕,快速規則的旋律可產生恐懼。音樂引發情緒的感覺有跨文化的效果,非洲民族能夠辨識西方音樂所展現的快樂、哀傷與恐懼,縱使從來沒有聽過。音樂和其他刺激一樣,會興奮腦中的情緒網路。好聽愉快的音樂會興奮腦中酬賞的神經回路(Blood & Zatore, 2001)。杏仁核受損病人無法辨識恐懼表情,也無法辨識音樂的恐懼(Gosselin et al., 2007)。
- 音樂知覺—和語文的關係:
- 顳葉平面(Planum temporale)是聽覺區的一部份,包含韋尼克區(Wernicke area)。
- 一般人65%左邊較右邊大,11%則相反。不對稱性在86%的嬰兒可以看出,但在左手者較不明顯。被認為和語言功能有關。
- 此一解剖不對稱性也存在猩猩腦中。其他語言相關區域亦然。猴子的左腦對於同類所發出的聲音也叫有反應。可能為語言先驅。
- 右腦的Planum temporale則與音樂有關。但是具有絕對音感者其左側的顳葉平面依然較大。 該區左右不對稱性如下:有絕對音感的音樂家>無絕對音感的音樂家>非音樂家